C형 간염 바이러스
"오늘의AI위키"의 AI를 통해 더욱 풍부하고 폭넓은 지식 경험을 누리세요.
1. 개요
C형 간염 바이러스(HCV)는 1989년 발견된 바이러스로, C형 간염의 원인이다. 지질 막과 두 개의 당단백질(E1, E2)로 구성된 외피, 정이십면체 코어와 RNA 유전체를 가지고 있다. HCV는 유전형에 따라 6가지 유형으로 분류되며, 유전자형에 따라 치료 반응과 치료 기간이 달라진다. 주로 혈액을 통해 전파되며, 현재 효과적인 백신은 없지만, 항바이러스제 개발로 치료 성공률이 높아졌다. 2020년에는 C형 간염 바이러스 발견에 기여한 하비 J. 알터, 마이클 호턴, 찰스 M. 라이스가 노벨 생리학·의학상을 수상했다.
더 읽어볼만한 페이지

2. 역사
C형 간염 바이러스의 기원을 밝히는 것은 어려웠지만, 1형과 4형은 공통 기원을 공유하는 것으로 보인다.[51] 베이즈 분석에 따르면 주요 유전자형은 약 300~400년 전에 공통 조상 바이러스에서 분화되었다.[52] 하위 유전자형은 약 200년 전에 주요 유전자형에서 분화되었다. 현존하는 모든 유전자형은 1형 1b 아형에서 진화한 것으로 보인다.
C형 간염 바이러스(HCV) 입자는 지름이 55~65nm인 지질 막 외피로 둘러싸여 있다.[10][11] 외피에는 두 개의 바이러스 외피 당단백질인 E1과 E2가 박혀 있는데,[12] 이들은 바이러스 부착 및 세포 내 진입에 관여한다.[10] 외피 안쪽에는 지름이 33~40nm인 정이십면체 코어가 있으며,[11] 이 코어 안에 바이러스의 RNA 물질이 들어있다.[10]
6형 계통에 대한 연구는 더 이른 진화 시점을 제시한다. 약 1,100~1,350년 현재로부터.[53] 추정 돌연변이율은 1.8 × 10−4였다. 실험 연구에서는 돌연변이율을 연간 위치당 2.5–2.9 × 10−3 염기 치환으로 추정했다.[54] 이 유전자형이 다른 유전자형의 조상일 수 있다.
유럽, 미국 및 일본의 분리주에 대한 연구에 따르면 1b형 유전자형의 기원은 약 1925년으로 추정된다.[55] 2a형과 3a형의 추정 기원 시점은 각각 1917년과 1943년이었다. 3형은 동남아시아에서 기원한 것으로 생각된다.[59]
이러한 다양한 국가의 연대기는 이 바이러스가 동남아시아에서 진화하여 서유럽의 상인에 의해 서아프리카로 확산되었을 수 있음을 시사한다.[60] 이후 국가의 쇄국 정책이 해제되면서 일본에 유입되었다. 1990년대에 혈액 제제에서 HCV 선별 검사가 시행된 후 확산 속도가 감소한 점을 감안할 때, 이전에는 수혈이 확산의 중요한 방법이었던 것으로 보인다.
1989년에 미국의 Chiron사 연구 그룹에 의해 비A형 비B형 간염 환자로부터 유전자 단편이 분리되었다.
2020년 10월 5일, C형 간염 바이러스의 특정에 기여한 발견을 한 공로로 하비 J. 올터, 마이클 호턴, 찰스 라이스가 노벨 생리학·의학상을 수상했다.[83]
3. 구조

3. 1. E1 및 E2 당단백질
E1과 E2는 C형 간염 바이러스(HCV)의 외피에 삽입될 때 공유 결합되어 있으며, 이황화 결합에 의해 안정화된다. E2는 구형이며 전자 현미경 이미지에 따르면 외피 막에서 6nm 정도 돌출되어 있는 것으로 보인다.[11]
이 당단백질들은 C형 간염 바이러스가 면역 체계와 상호 작용하는 데 중요한 역할을 한다. E2 당단백질에는 초변이 영역, 즉 초변이 영역 1(HVR1)이 존재한다.[10] HVR1은 유연하며 주변 분자에 매우 접근하기 쉽다.[13] HVR1은 E2가 바이러스를 면역 체계로부터 보호하는 데 도움을 주는데, 이는 CD81이 바이러스의 해당 수용체에 결합하는 것을 막기 때문이다.[13] 또한, E2는 E1을 면역 체계로부터 보호할 수 있다.[13] HVR1은 아미노산 서열에서 매우 가변적이지만, 이 영역은 많은 E2 당단백질에서 유사한 화학적, 물리적 및 구조적 특성을 갖는다.[14]
4. 유전체
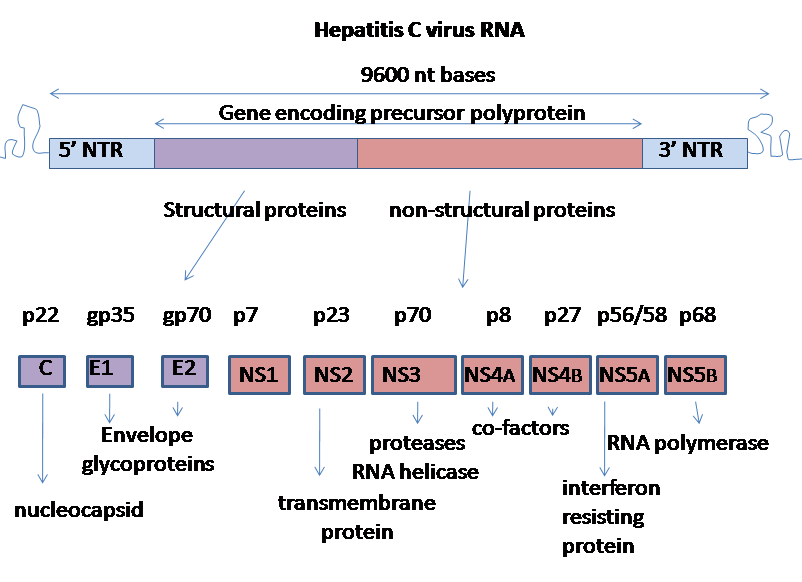

C형 간염 바이러스는 플러스 가닥 단일 가닥 RNA 유전체를 가지고 있다. 유전체는 길이가 9,600 뉴클레오타이드 염기쌍인 단일 개방형 읽기 틀로 구성된다.[15] 이 단일 개방형 읽기 틀은 번역되어 단일 단백질 산물을 생성하며, 이 단백질은 더 작고 활성적인 단백질을 생성하기 위해 추가로 처리된다.
RNA의 5' 및 3' 말단에는 단백질로 번역되지 않지만 바이러스 RNA의 번역 및 복제에 중요한 비번역 영역(UTR)이 있다. 5' UTR에는 약 3,000개의 아미노산을 포함하는 매우 긴 단백질의 번역을 시작하는 리보솜 결합 부위[16] 또는 내부 리보솜 진입 부위(IRES)가 있다. HCV IRES의 핵심 도메인은 예측된 유사매듭 내에 통합된 사방 나선 홀리데이 접합을 포함한다.[17] 이 핵심 도메인의 컨포메이션은 40S 리보솜 소단위체에 위치하기 위한 개방형 읽기 틀의 방향을 제한한다. 큰 전구 단백질은 나중에 세포 및 바이러스 단백질 분해 효소에 의해 절단되어 숙주 세포 내에서 바이러스 복제를 허용하거나 성숙한 바이러스 입자로 조립되는 10개의 더 작은 단백질로 변환된다.[18]
C형 간염 바이러스에 의해 생성되는 구조 단백질에는 코어 단백질, E1 및 E2가 포함되며, 비구조 단백질에는 NS2, NS3, NS4A, NS4B, NS5A, 및 NS5B가 포함된다. 단일 가닥 RNA는 약 3,010개의 아미노산으로 구성된 폴리프로테인을 코딩할 수 있는 열린 읽기 틀(ORF)을 가지고 있으며, 이 폴리프로테인, 즉 전구체 단백질에 세포의 시그날라제와 바이러스 자체가 코딩하는 두 종류의 프로테아제가 작용하여 바이러스 입자를 형성하는 구조 단백질과 바이러스 입자에 포함되지 않는 비구조 단백질이 생산된다. 게놈의 5' 말단에는 바이러스 단백질의 번역 조절에 관여하는 영역이 존재하며, 이는 다양성이 높은 게놈 배열 중에 있으며 바이러스 클론 간에 가장 잘 보존되어 있어 유전자 검출에 이용된다.[84]
5. 분자생물학
C형 간염 바이러스는 플러스 가닥 단일 가닥 RNA 게놈을 가진다. 게놈은 9,600 뉴클레오타이드 염기쌍 길이의 단일 개방형 읽기 틀로 구성된다.[15] 이 단일 개방형 읽기 틀은 번역되어 하나의 단백질을 생성하며, 이 단백질은 추가 처리 과정을 거쳐 더 작고 활성적인 여러 단백질로 나뉜다.
RNA의 5' 말단과 3' 말단에는 단백질로 번역되지 않지만 바이러스 RNA의 번역과 복제에 중요한 비번역 영역(UTR)이 존재한다. 5' UTR에는 리보솜 결합 부위[16] 또는 내부 리보솜 진입 부위(IRES)가 있어, 약 3,000개의 아미노산을 포함하는 매우 긴 단백질의 번역을 시작한다. HCV IRES의 핵심 도메인은 유사매듭 내에 통합된 사방 나선 홀리데이 접합을 포함한다.[17] 이 도메인의 구조는 40S 리보솜 소단위체에 결합하여 번역을 시작할 수 있도록 돕는다. 큰 전구 단백질은 세포와 바이러스의 단백질 분해 효소에 의해 절단되어 10개의 작은 단백질로 나뉘며, 이들은 숙주 세포 내에서 바이러스 복제를 돕거나 성숙한 바이러스 입자를 구성한다.[18]
C형 간염 바이러스 유전자는 다음과 같은 순서로 배열되어 있다: N 말단-코어-외피(E1)–E2–p7-비구조 단백질 2(NS2)–NS3–NS4A–NS4B–NS5A–NS5B–C 말단.
NS2/NS3 접합부는 NS2의 금속 의존적 자가 촉매 단백질분해효소와 NS3의 N-말단에 의해 절단된다. NS4B는 막성 웹 형성에 관여한다. 캡시드 유전자에서 +1 프레임 시프트에 의해 암호화되는 11번째 단백질도 존재한다.[27][28] 이 단백질은 항원성을 갖지만, 기능은 알려져 있지 않다.
6. 복제
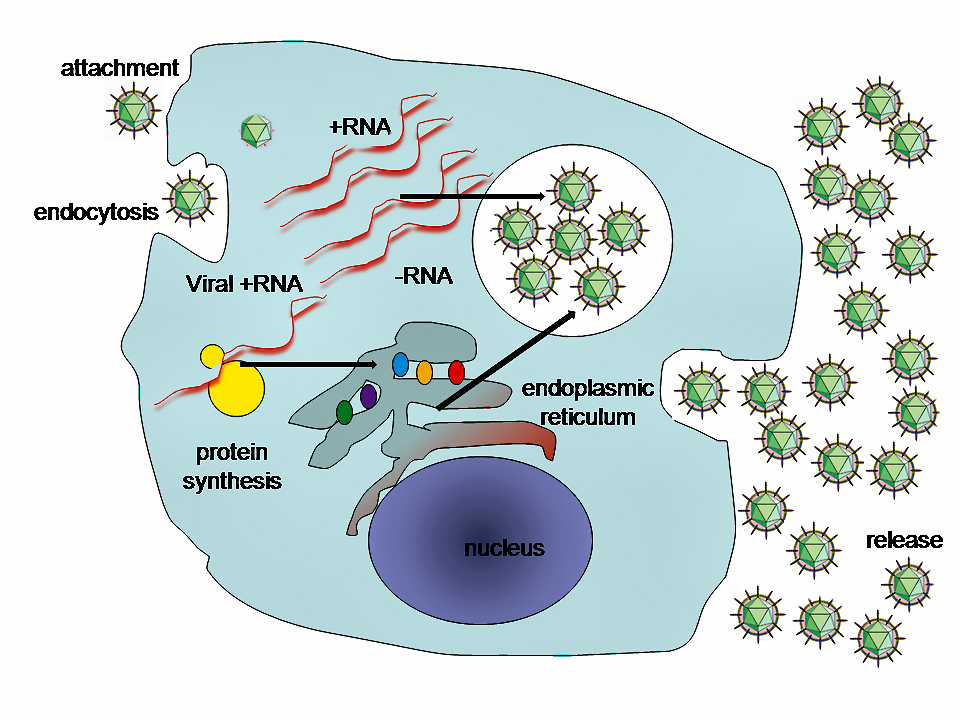
C형 간염 바이러스(HCV)는 주로 간세포에서 복제되며, 감염된 각 세포는 매일 약 50개의 바이러스 입자(비리온)를 생성하여 하루에 총 1조 개의 비리온이 생성되는 것으로 추정된다.[10] HCV는 말초 혈액 단핵 세포에서도 복제될 수 있어 만성 HCV 감염 환자에게서 높은 수준의 면역 장애가 나타날 수 있다.
HCV는 여러 유전자형을 가지며, 바이러스의 RNA 의존성 RNA 중합 효소의 높은 오류율로 인해 빠르게 변이한다. 이러한 변이율 때문에 다양한 바이러스 변종이 생성되어 준 종으로 간주된다.[29]
숙주 세포로의 진입은 바이러스의 당단백질과 CD81, LDL 수용체, SR-BI, DC-SIGN, 클라우딘-1, 오클루딘 등 세포 표면 분자와의 복잡한 상호 작용을 통해 일어난다.[30][31] HCV 외피는 초저밀도 지단백질(VLDL) 및 저밀도 지단백질(LDL)과 유사하여 아포지단백질과 연관될 수 있다. 최근 연구에 따르면 아포지단백질은 청소 수용체 B1(SR-B1)과 상호 작용하여 바이러스 주변 지질을 제거하고 HVR1 접촉을 용이하게 한다. 타이트 접합 단백질인 클라우딘 1과 CD81은 복합체를 형성하여 HCV 감염을 준비한다. 면역 체계가 작동하면 대식세포는 감염된 간세포 주변의 TNF-α 양을 증가시켜 타이트 접합 복합체인 오클루딘의 기저 외측 막으로의 이동을 유발한다.[10]
이러한 상호 작용으로 바이러스 입자는 세포내이입되고, 이 과정은 클라트린 단백질에 의해 지원된다. 초기 엔도솜 내에서 엔도솜과 바이러스 외피가 융합되면 RNA가 세포질로 방출된다.[10]
HCV는 세포 내 기계의 일부를 이용하여 복제한다.[32] HCV 유전체는 약 3,011개의 아미노산으로 구성된 단일 단백질로 번역된다. 이 폴리단백질은 바이러스 및 세포 프로테아제에 의해 단백질 분해적으로 처리되어 3개의 구조 단백질과 7개의 비구조 단백질(NS)을 생성한다. Core 영역에서 프레임 시프트가 발생하면 대체 리딩 프레임 단백질(ARFP)이 생성될 수 있다.[33] HCV는 NS2 시스테인 자가 프로테아제와 NS3-4A 세린 프로테아제라는 두 개의 프로테아제를 암호화한다. NS 단백질은 바이러스 유전체를 재배열된 세포질 막과 관련된 RNA 복제 복합체로 모집한다. RNA 복제는 바이러스 RNA 의존성 RNA 중합 효소 NS5B를 통해 일어나며, 음성 가닥 RNA 중간체를 생성한다. 음성 가닥 RNA는 새로운 양성 가닥 바이러스 유전체 생산을 위한 주형 역할을 한다. 생성된 유전체는 번역, 추가 복제, 또는 새로운 바이러스 입자 내에서 패키징될 수 있다.
바이러스는 세포 내 지질 막, 특히 소포체에서 복제된다.[34] 소포체는 '막질 웹'이라는 독특한 모양의 막 구조로 변형되며, 이는 바이러스 단백질 NS4B의 발현에 의해 유도될 수 있다.[35] 코어 단백질은 지질 방울과 연관되어 미세 소관과 다이닌을 활용하여 핵주위로 이동한다.[36] 간세포에서의 방출은 VLDL 분비 경로와 관련될 수 있다.[37] 또는, 바이러스 입자가 수송에 필요한 엔도솜 소팅 복합체(ESCRT) 경로를 통해 소포체에서 분비될 수 있다는 가설도 있다.[10]
7. 유전자형
C형 간염 바이러스(HCV)는 유전적 차이에 따라 6개의 주요 유전자형(1~6)으로 분류되며, 각 유전자형 안에는 여러 아형(소문자로 표시)이 존재한다.[38][39] 유전자형은 전체 유전체의 뉴클레오티드 배열에서 30~35% 정도 차이가 난다.[40] 1a형과 1b형은 전 세계적으로 가장 흔하며, 전체 C형 간염의 60%를 차지한다.
C형 간염 바이러스 유전자형의 기원을 밝히는 것은 쉽지 않지만, 1형과 4형은 공통 조상을 공유하는 것으로 추정된다.[51] 베이즈 정리에 따르면 주요 유전자형들은 약 300~400년 전에 공통 조상 바이러스로부터 분화되었고,[52] 하위 유전자형들은 약 200년 전에 분화되었다. 6형은 약 1,100~1,350년 전에 분화되어 다른 유전자형의 조상일 가능성이 제기되기도 한다.[53]
유럽, 미국, 일본 등지의 연구에 따르면 1b형은 1925년경, 2a형과 3a형은 각각 1917년과 1943년에 기원한 것으로 추정된다.[55] 3형은 동남아시아에서 기원한 것으로 알려져 있다.[59]
C형 간염 바이러스는 지역별로 유행하는 유전자형에 차이가 있다. 미국에서는 1a형, 유럽에서는 1a형과 3a형이 흔하다. 일본에서는 1b형이 70%로 가장 많고, 2a형과 2b형이 그 뒤를 잇는다. 한국에서는 1b형이 가장 흔하며, 다음으로 2a형, 2b형 순으로 발견된다.
일본에서는 치료 효과와 관련하여 유전자형을 2개의 혈청형으로 분류하기도 한다.
- 1군 (Group1): 주로 1a형, 1b형, 1c형
- 2군 (Group2): 주로 2a형, 2b형, 2c형
C형 간염 바이러스에 감염되면 거의 항상 항체가 생성되므로, 항체 검사는 진단에 매우 중요하다.
7. 1. 임상적 중요성
유전자형은 인터페론 기반 치료에 대한 잠재적 반응과 치료 기간을 결정하는 데 중요하게 작용한다. 유전자형 1형과 4형은 다른 유전자형(2, 3, 5, 6형)보다 인터페론 기반 치료에 덜 반응한다.[41] 표준 인터페론 기반 치료 기간은 1형과 4형의 경우 48주이고, 2형과 3형의 경우 24주면 완료된다. 지속적인 바이러스 반응은 1형의 경우 70%, 2형과 3형의 경우 ~90%, 4형의 경우 ~65%, 6형의 경우 ~80%에서 발생한다.[42] 아프리카계 사람들은 1형 또는 4형 유전자형에 감염된 경우 치료에 반응할 가능성이 훨씬 낮다.[43]한 유전자형에 감염되어도 다른 유전자형에 대한 면역력이 생기지 않으며, 두 균주의 동시 감염도 가능하다. 이러한 경우 대부분 한 균주가 짧은 시간 안에 다른 균주보다 우위를 차지한다. 이 발견은 치료에 유용할 수 있으며, 약물에 반응하지 않는 균주를 치료하기 쉬운 다른 균주로 대체하는 데 활용될 수 있다.[46]
8. 역학
C형 간염 바이러스는 주로 혈액 매개 바이러스이며, 성적 또는 수직 감염 위험은 매우 낮다.[48] 이러한 전파 방식으로 인해 위험이 높은 주요 집단은 정맥 주사 약물 사용자(IDU), 혈액 제제 수혜자, 때로는 혈액 투석 환자이다. C형 간염의 흔한 전파 환경은 또한 병원 내 감염 (원내 감염)으로, 진료소에서 위생 및 멸균 관행이 제대로 준수되지 않을 때 발생한다.[49] 할례, 여성 할례, 의례적 흉터, 전통 문신 및 침술을 포함하여 여러 문화적 또는 의례적 관행이 HCV의 잠재적인 역사적 전파 방식으로 제안되었다.[48] 또한, 사람에게서 HCV가 매우 오랫동안 지속된다는 점을 감안할 때, 물린 곤충을 통한 매우 낮고 감지할 수 없는 기계적 전파율조차 사람들이 많은 수의 곤충 물림을 받는 열대 지방에서 풍토성 감염을 유지하기에 충분할 수 있다고 주장되어 왔다.[50]
바이러스의 전파는 수혈이나 의료 관계자의 바늘 찔림 사고[85] 등 혈액을 매개로 한 것 및 성행위에 의한 감염[85], 특히 고령 부부 사이의 감염이 우려된다.[86]
9. 면역 회피
HCV는 간세포와 일부 림프구를 표적 세포로 하며, 숙주의 면역 기구와 인터페론으로부터 탈출하여 지속적인 감염을 일으킨다. 감염된 세포나 수지상 세포가 생산하는 인터페론(IFN)은 C형 간염 바이러스(HCV) 제거에 작용하며, NK 세포도 수지상 세포에 의해 활성화되어 바이러스 제거에 기여한다. 그러나 HCV는 NS3 등을 통해 인터페론 신호 전달을 저해하고, E2에 의해 NK 세포의 기능을 저하시켜 면역계로부터 회피한다.[89]
10. 치료
C형 간염은 효과적인 백신은 없지만, 항바이러스제로 치료가 가능하다. 2015년 7월에 승인된 레디파스비르와 같은 항바이러스제는 96~100%의 환자에게서 바이러스를 제거할 수 있는 높은 치료 효과를 보인다.[93]
11. 백신
A형과 B형 간염과 달리, 현재 C형 간염을 예방할 수 있는 백신은 존재하지 않는다.[96] 항바이러스제인 레디파스비르가 2015년 7월에 승인되어 C형 간염 바이러스를 96~100%의 사람에게서 제거하는 것이 가능해졌지만, 효과적인 백신은 아직 없다.[93]
12. 현재 연구
C형 간염 바이러스 연구는 바이러스의 좁은 숙주 범위 때문에 어려움을 겪어왔다.[62] 그러나 최근 레플리콘 시스템이 개발되어 연구에 활용되고 있다.[63] C형 간염 바이러스는 대부분의 RNA 바이러스처럼 바이러스 유사종으로 존재하기 때문에, 단일 균주나 수용체 유형을 분리하여 연구하기가 매우 어렵다.[64][65]
현재 연구는 바이러스의 단백질 분해 효소, RNA 중합 효소 및 기타 비구조 유전자의 소분자 억제제에 집중되어 있다. 머크사의 보세프레비르(boceprevir)와 버텍스 파마슈티컬스의 인시베크(Incivek)는 NS3 단백질 분해 효소 억제제로, 2011년에 사용이 승인되었다.[66]
낮은 비타민 D 수치와 치료 반응 불량 사이의 연관성이 보고되었으며,[67][68][69][70] ''시험관 내'' 연구에서 비타민 D가 바이러스 복제를 감소시킬 수 있다는 결과가 나왔다.[71] 임상 시험 결과는 아직 기다려야 하지만,[74][75] 표준 치료 외에 비타민 D 보충이 치료 반응을 향상시키는 데 중요하다고 제안되었다.[76]
나린제닌은 자몽 등에서 발견되는 플라보노이드로, 세포 내 감염성 바이러스 입자의 조립을 차단하는 것으로 나타났다.[76]
뉴클레오사이드 및 뉴클레오타이드 유사체 억제제, RNA 의존성 RNA 중합 효소의 비-뉴클레오사이드 억제제, NSP5A 억제제, 사이클로필린 억제제, 실리비닌과 같은 숙주 표적 화합물도 연구 중이다.[77]
소포스부비르(sofosbuvir)는 만성 C형 간염 감염 치료제로 2013년 FDA 승인을 받았다. 이는 인터페론 병용 투여 없이 특정 유형의 C형 간염 감염을 치료하는 안전성과 효능을 입증한 최초의 약물로 보고되었다.[78] 시메프레비르(simeprevir)는 페그인터페론-알파(peginterferon-alfa) 및 리바비린(ribavirin)과 함께 사용하도록 FDA 승인을 받았으며,[79] 일본에서도 만성 C형 간염 감염, 유전자형 1의 치료를 위해 승인되었다.[80]
옥시마트린(oxymatrine)은 아시아에서 발견되는 뿌리 추출물로, 세포 배양 및 동물 연구에서 C형 간염 바이러스에 대한 항바이러스 활성이 있다고 보고되었다. 소규모 인간 임상 시험에서 유익한 결과와 심각한 부작용이 없는 것으로 나타났지만, 일반적인 결론을 내리기에는 규모가 작았다.[76]
2020년 10월 5일, 하비 J. 알터(Harvey J. Alter), 마이클 호턴(Michael Houghton), 찰스 M. 라이스(Charles M. Rice)가 C형 간염 바이러스의 발견으로 2020년 노벨 생리학·의학상을 수상했다.[81]
참조
[1]
NCBI taxid
Hepacivirus C
2022-01-03
[2]
웹사이트
Create 13 new species in the genus Hepacivirusand rename 1 species (family Flaviviridae)
https://ictv.global/[...]
2016-06-23
[3]
웹사이트
Genus: Hepacivirus
https://web.archive.[...]
International Committee on Taxonomy of Viruses
2018-07
[4]
학술지
HCV syndrome: A constellation of organ- and non-organ specific autoimmune disorders, B-cell non-Hodgkin's lymphoma, and cancer
[5]
학술지
Mechanisms of HCV-induced liver cancer: what did we learn from in vitro and animal studies?
[6]
학술지
Characterization of a canine homolog of hepatitis C virus
[7]
학술지
Serology-enabled discovery of genetically diverse hepaciviruses in a new host
2012-06
[8]
학술지
Bats are a major natural reservoir for hepaciviruses and pegiviruses
2013-05
[9]
학술지
Identification of rodent homologs of hepatitis C virus and pegiviruses
[10]
학술지
Virology and cell biology of the hepatitis C virus life cycle – An update
[11]
학술지
Morphology of hepatitis C and hepatitis B virus particles as detected by immunogold electron microscopy
2006-06
[12]
학술지
Topology of hepatitis C virus envelope glycoproteins
[13]
학술지
A Biologically-validated HCV E1E2 Heterodimer Structural Model
2017-03-16
[14]
학술지
The Hypervariable Region 1 of the E2 Glycoprotein of Hepatitis C Virus Binds to Glycosaminoglycans, but This Binding Does Not Lead to Infection in a Pseudotype System
2004-05
[15]
학술지
Genome of human hepatitis C virus (HCV): gene organization, sequence diversity, and variation
[16]
학술지
Hepatitis C IRES: translating translation into a therapeutic target
[17]
학술지
Crystal structure of the HCV IRES central domain reveals strategy for start-codon positioning
2011-10
[18]
학술지
Hepatitis C virus proteins
[19]
학술지
Molecular virology of the hepatitis C virus
[20]
학술지
Intrinsically unstructured domain 3 of hepatitis C Virus NS5A forms a "fuzzy complex" with VAPB-MSP domain which carries ALS-causing mutations
[21]
학술지
Assembly, purification, and pre-steady-state kinetic analysis of active RNA-dependent RNA polymerase elongation complex
[22]
학술지
Replication of hepatitis C virus
2007-06
[23]
학술지
Ligand-induced changes in hepatitis C virus NS5B polymerase structure
2010-11
[24]
학술지
Crystal structures of the RNA-dependent RNA polymerase genotype 2a of hepatitis C virus reveal two conformations and suggest mechanisms of inhibition by non-nucleoside inhibitors
2005-05
[25]
학술지
Substrate complexes of hepatitis C virus RNA polymerase (HC-J4): structural evidence for nucleotide import and de-novo initiation
2003-02
[26]
학술지
Non-nucleoside inhibitors binding to hepatitis C virus NS5B polymerase reveal a novel mechanism of inhibition
2006-08
[27]
학술지
Evidence for a new hepatitis C virus antigen encoded in an overlapping reading frame
[28]
학술지
Expression and characterization of Escherichia coli derived hepatitis C virus ARFP/F protein
[29]
학술지
Replication of hepatitis C virus
https://web.archive.[...]
2010-07-16
[30]
학술지
Hepatitis C virus entry: molecular mechanisms and targets for antiviral therapy
[31]
학술지
Splicing diversity of the human OCLN gene and its biological significance for hepatitis C virus entry
[32]
학술지
Unravelling hepatitis C virus replication from genome to function
[33]
학술지
The Hepatitis C Virus Alternate Reading Frame (ARF) and Its Family of Novel Products: The Alternate Reading Frame Protein/F-Protein, the Double-Frameshift Protein, and Others
[34]
학술지
Interaction of hepatitis C virus proteins with host cell membranes and lipids
[35]
학술지
Expression of hepatitis C virus proteins induces distinct membrane alterations including a candidate viral replication complex
[36]
논문
Hepatitis C virus core protein induces lipid droplet redistribution in a microtubule- and dynein-dependent manner
[37]
논문
Hepatitis C virus hijacks host lipid metabolism
[38]
논문
Classification of hepatitis C virus into six major genotypes and a series of subtypes by phylogenetic analysis of the NS-5 region
https://www.microbio[...]
1993-11-01
[39]
논문
An updated analysis of hepatitis C virus genotypes and subtypes based on the complete coding region
2011-10-09
[40]
논문
New hepatitis C virus (HCV) genotyping system that allows for identification of HCV genotypes 1a, 1b, 2a, 2b, 3a, 3b, 4, 5a, and 6a
[41]
논문
Consensus proposals for a unified system of nomenclature of hepatitis C virus genotypes
[42]
논문
Treatment of chronic hepatitis C in Asia: when East meets West
[43]
논문
Peginterferon alfa-2b and ribavirin for the treatment of chronic hepatitis C in blacks and non-Hispanic whites.
2004-01-01
[44]
논문
Genetic variation in IL28B predicts hepatitis C treatment-induced viral clearance
2009-01-01
[45]
논문
Viral evolution explains the associations among hepatitis C virus genotype, clinical outcomes, and human genetic variation
2013-01-01
[46]
논문
Exposure of hepatitis C virus (HCV) RNA-positive recipients to HCV RNA-positive blood donors results in rapid predominance of a single donor strain and exclusion and/or suppression of the recipient strain
[47]
논문
Recombination in hepatitis C virus
2011-10-01
[48]
논문
Global epidemiology of hepatitis C virus infection
http://www.thelancet[...]
2005-09-01
[49]
논문
HCV routes of transmission: what goes around comes around
2011-11-01
[50]
논문
Investigating the endemic transmission of the hepatitis C virus
2007-07-01
[51]
논문
Hepatitis C virus evolutionary patterns studied through analysis of full-genome sequences
[52]
논문
NS4A protein as a marker of HCV history suggests that different HCV genotypes originally evolved from genotype 1b
[53]
논문
Genetic history of hepatitis C virus in East Asia
[54]
논문
Study of multiple genetic variations caused by persistent hepatitis C virus replication in long-term cell culture
[55]
논문
Investigation of the pattern of diversity of hepatitis C virus in relation to times of transmission
[56]
논문
The global spread of hepatitis C virus 1a and 1b: a phylodynamic and phylogeographic analysis
2009-12-01
[57]
논문
Phylogeography and molecular epidemiology of hepatitis C virus genotype 2 in Africa
2009-09-01
[58]
논문
Colonial History and Contemporary Transmission Shape the Genetic Diversity of Hepatitis C Virus Genotype 2 in Amsterdam
2012-01-01
[59]
논문
Genetic diversity and evolution of hepatitis C virus—15 years on
2004-11-01
[60]
논문
Reconstructing the origins of human hepatitis viruses
[61]
논문
A new insight into hepatitis C vaccine development
[62]
논문
Host genetic determinants of spontaneous hepatitis C clearance
[63]
논문
Hepatitis C virus virology and new treatment targets
2009-04-01
[64]
논문
The way forward in HCV treatment—finding the right path
2007-12-01
[65]
논문
Beclabuvir in combination with asunaprevir and daclatasvir for hepatitis C virus genotype 1 infection: A systematic review and meta-analysis
2018-05-01
[66]
웹사이트
FDA approves Victrelis for Hepatitis C
https://www.fda.gov/[...]
FDA
2011-05-13
[67]
논문
Classical and emerging roles of vitamin d in hepatitis C virus infection
[68]
논문
Vitamin D deficiency and a CYP27B1-1260 promoter polymorphism are associated with chronic hepatitis C and poor response to interferon-alfa based therapy
https://www.journal-[...]
[69]
논문
The vitamin D receptor gene bAt (CCA) haplotype impairs the response to pegylated-interferon/ribavirin-based therapy in chronic hepatitis C patients
[70]
논문
Complementary role of vitamin D deficiency and the interleukin-28B rs12979860 C/T polymorphism in predicting antiviral response in chronic hepatitis C
[71]
논문
Vitamin D: an innate antiviral agent suppressing hepatitis C virus in human hepatocytes
[72]
논문
Vitamin D supplementation improves sustained virologic response in chronic hepatitis C (genotype 1)-naïve patients
[73]
논문
Vitamin D supplementation improves response to antiviral treatment for recurrent hepatitis C
[74]
논문
Review article: the extra-skeletal effects of vitamin D in chronic hepatitis C infection
2012-03
[75]
논문
Diagnostic and therapeutical role of vitamin D in chronic hepatitis C virus infection
[76]
논문
Alternative medications in Hepatitis C infection
2014-01-27
[77]
논문
Antiviral strategies in hepatitis C virus infection
[78]
웹사이트
Press announcement, FDA, December 6 2013
https://www.fda.gov/[...]
[79]
웹사이트
FDA approves new treatment for hepatitis C virus
https://www.fda.gov/[...]
Food and Drug Administration
2013-11-22
[80]
웹사이트
Medivir: Simeprevir has been approved in Japan for the treatment of genotype 1 chronic hepatitis C infection
https://www.wsj.com/[...]
The Wall Street Journal
2013-09-27
[81]
웹사이트
The Nobel Prize in Physiology or Medicine 2020
https://www.nobelpri[...]
[82]
간행물
ウイルス肝炎の最新治療と今後の展望 〜超音波検査に必要な最新情報〜
https://doi.org/10.1[...]
[83]
웹사이트
ノーベル医学生理学賞、ハービー・J・オルター氏ら3氏 C型肝炎ウイルス発見
https://www.cnn.co.j[...]
CNN
2020-10-05
[84]
웹사이트
NIID 国立感染症研究所『C型肝炎とは』
https://www.niid.go.[...]
2019-12-18
[85]
논문
医療従事者の針刺し事故によるC型肝炎ウイルス感染について
https://doi.org/10.1[...]
[86]
논문
結婚40年後の配偶者間性行為感染が疑われたC型急性肝炎の1例
https://doi.org/10.2[...]
[87]
뉴스
C型肝炎ウイルス 中性脂肪利用し増殖
http://www.yomiuri.c[...]
読売新聞
2007-08-27
[88]
간행물
The lipid droplet is an important organelle for hepatitis C virus production
http://www.nature.co[...]
[89]
웹사이트
HCV蛋白の構造と機能
http://www0.nih.go.j[...]
国立感染症研究所
[90]
논문
本邦のC型慢性肝疾患におけるクリオグロブリン血症についての検討
https://doi.org/10.1[...]
[91]
논문
C型肝炎ウイルスと肝外病変
https://doi.org/10.1[...]
[92]
논문
C型肝炎ウイルスによる糖代謝異常
https://doi.org/10.2[...]
[93]
웹사이트
ハーボニー:著効率100%のC型肝炎治療薬
https://medical.nikk[...]
日経メディカルオンライン
2015-09-19
[94]
저널
HCV syndrome: A constellation of organ- and non-organ specific autoimmune disorders, B-cell non-Hodgkin’s lymphoma, and cancer
[95]
저널
Mechanisms of HCV-induced liver cancer: what did we learn from in vitro and animal studies?
[96]
저널
A new insight into hepatitis C vaccine development
본 사이트는 AI가 위키백과와 뉴스 기사,정부 간행물,학술 논문등을 바탕으로 정보를 가공하여 제공하는 백과사전형 서비스입니다.
모든 문서는 AI에 의해 자동 생성되며, CC BY-SA 4.0 라이선스에 따라 이용할 수 있습니다.
하지만, 위키백과나 뉴스 기사 자체에 오류, 부정확한 정보, 또는 가짜 뉴스가 포함될 수 있으며, AI는 이러한 내용을 완벽하게 걸러내지 못할 수 있습니다.
따라서 제공되는 정보에 일부 오류나 편향이 있을 수 있으므로, 중요한 정보는 반드시 다른 출처를 통해 교차 검증하시기 바랍니다.
문의하기 : help@durumis.com